Bat echolocation has fascinated scientists for decades (Griffin 1958), creating a bulk of literature (reviewed in Jones & Teeling 2006, Jones & Holderied 2007) that covers various aspects ranging from neurophysiological control of sound production (e.g. Suthers & Fattu 1973) to the auditory scene analysis applied during sound perception (e.g. Moss & Surlyyke 2001). In recent years, scientists have become increasingly aware that bat echolocation calls serve communicative functions (reviewed in Fenton 2003, Jones & Siemers 2010) in addition to their primary function of facilitating orientation and foraging.
Communicative function of echolocation
A growing body of evidence indicates that echolocation calls have the potential to transmit information about lucrative foraging patches (e.g. Gillam 2007), suitable roost sites (e.g. Ruczynski et al. 2007), species and individual identity (e.g. Russo et al. 2007), age (e.g. Jones & Ransome 1993), sex (e.g. Jones et al. 1992) and social group affiliation (e.g. Jameson & Hare 2009) of the calling bat. Several laboratory studies provide experimental evidence to what extent the transmitted information is actually exploited by eavesdroppers (Kazial et al. 2004, Yovel et al. 2009, Schuchmann & Siemers 2010, Voigt-Heuke et al. 2010).
Recently, we contributed to this field of research by providing conclusive experimental evidence that free-living male greater sac-winged bats (Saccopteryx bilineata) can discriminate the sex of conspecifics based on echolocation call parameters alone (Knörnschild et al. 2012).
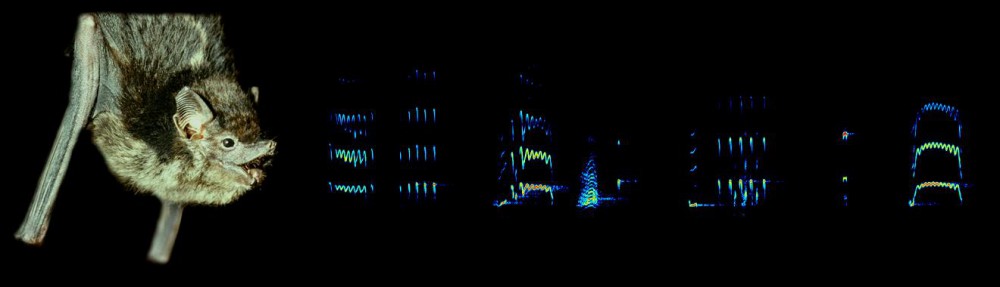
 Echolocation calls of Saccopteryx bilineata (© Jung).
Echolocation calls of Saccopteryx bilineata (© Jung).
Following up on this discovery, my colleague Kirsten Jung and I are currently investigating to which extent the acoustic variation in echolocation calls of Saccopteryx bilineata is influenced by natural and sexual selection. To elucidate this, we are analyzing echolocation calls from different Saccopteryx bilineata populations in Costa Rica. We are expecting that the sex-specific signature in echolocation calls will prevail over acoustic differences caused by the various natural habitats in which our study populations live.
Griffin. 1958. Listening in the Dark: the Acoustic Orientation of Bats and Man. Yale University Press
Gillam. 2007. Can J Zool 85:795-801
Jones et al. 1992. Mammalia 56:189-193
Jones & Ransome. 1993. Proc R Soc B 252:125-128
Jones & Teeling. 2006. Trends Ecol Evol 21:149-156
Jones & Holderied. 2007. Proc R Soc B 274:905-912
Jameson & Hare. 2009. Acta Chiropterologica 11:163-172
Jones & Siemers. 2010. J Comp Physiol A 197:447-457
Kazial & Masters. 2004. Anim Behav 67:855-863
Knörnschild et al. 2012. Proc R Soc B 279:4827-4835
Moss & Surlykke. 2001. JASA 110:2207-2225
Ruczynski et al. 2007. J Exp Biol 210:3607-3615
Russo et al. 2007. J Biogeo 34:2129-2138
Schuchmann & Siemers. 2010. Am Nat 176:72–82
Suthers & Fattu. 1973. Am Zool 13:1215-1226
Voigt-Heucke et al. 2010. Anim Behav 80:59-67
Yovel et al. 2009. PLOS Comp Biol 5:e1000400